The Biology of the Goat
Life Cycle of the CAE virus
The CAE virus infects monocytes and macrophages. Monocytes are a type of white blood cell which circulate in the blood. The CAE virus cannot replicate inside a monocyte but its genome (called a provirus) is carried to all tissues of the body without being detected by the immune system. When a monocyte migrates to body tissues it matures into a macrophage. When this happens, factors are expressed by the cell which the provirus requires for replication. The macrophage then becomes a factory for production of infectious viruses which attack more monocytes and mature macrophages.
There are a few studies which indicate that the CAE virus may also be able to infect other types of cells. However, since the expression of virus proteins in these cases is restricted and the number of cells infected is very low this is probably not a common source of CAE infection.
The first step of infection: gaining access to the cell
The virus first binds to a surface molecule of a monocyte/macrophage using the envelope protein. Cells have many kinds of proteins on their surface which they use for their normal activities. Viruses exploit these receptors to gain entrance to the cell. The CAE virus will know that it is near a monocyte or macrophage because the envelope protein on the surface of the virus will fit only with a cell surface protein that exists only on goat (and sheep) monocyte/ macrophage cells. The envelope protein on the virus is a key that must fit exactly to a cellular lock to gain entrance to the cell. The reason that the CAE virus cannot infect humans is because human cells do not have the correct receptor on the cell surface.The animation below shows the steps of binding of the Human Immunodeficiency Virus (HIV) to the cell surface protein. HIV, also a lentivirus, is similar to the CAE virus. While the exact details of receptor binding in CAEV is not known it probably employs a similar strategy as HIV. Once the receptor binds to the first protein, this causes the SU portion of the envelope protein to go through a structural change, revealing another site that can bind to a second cell receptor. This brings the membrane of the virus even closer to the cell surface. Note that the second receptor site on the virus protein is hidden until it is needed in the binding process. This is one way that the virus is able to evade the immune response.
Second step of infection: converting RNA into DNA

Fusion of the membranes of the virus and host cell allow the the capsid to enter the cell. The capsid undergoes a structural change which partially opens it up. This allows access to required cell components. The capsid is now called a preintegration complex or PIC.
Inside the PIC reverse transcriptase converts the single strand of RNA into double-strand complementary DNA (cDNA). This is a complex, error prone process which leaves mutations in the newly created DNA. This is the reason that there are many variant forms of the CAE retrovirus.
Third step of infection: integration of DNA into cell chromosome
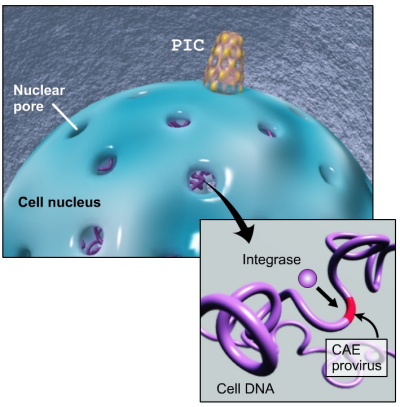 The PIC migrates to the nucleus of the cell carrying the cDNA and another of the proteins the virus brought along called integrase. cDNA and integrase is inserted into the nucleus through a nuclear pore.
The PIC migrates to the nucleus of the cell carrying the cDNA and another of the proteins the virus brought along called integrase. cDNA and integrase is inserted into the nucleus through a nuclear pore.Integrase clips off a base pair at the end of the viral cDNA, cuts open the strands of host DNA, then binds the ends of the viral DNA into the host DNA. Repair proteins supplied by the cell come along and clean up the loose ends.
The genome of the CAE virus is now a permanent part of the goat's monocyte/macrophage DNA. At this stage it's called a provirus.
Fourth step of infection: replication of new viruses
 The proviral DNA is transcribed into RNA which is transferred from the nucleus to the cytoplasm.
The proviral DNA is transcribed into RNA which is transferred from the nucleus to the cytoplasm.
Viral RNA is either used as a template to make viral proteins with the help of cell machinery or is packaged into the newly made viral capsid. Viral mRNA is transcribed into envelope protein which is transferred to the cell membrane. Capsid proteins, reverse transcriptase, integrase, protease and a few other proteins are also created from transcription of RNA.
Finally, two copies of RNA plus proteins are packaged into the newly created capsid and the capsid buds out from the cell through the modified membrane. The membrane of the virus came from the cell so this is why the CAE retrovirus is called an enveloped virus.
The new, free virus must go through a maturation period before it can become infective which among other things prevents it from immediately reinfecting the cell that produced it.
The genome of the CAE virus

This is a graphic representation of the genome of the CAE virus. The genome shown in yellow has sequences on each end called LTR for Long Terminal Repeats. LTRs, present in all retroviruses, are important in establishing the provirus in the host cell DNA and contain signals that are involved in expressing the viral genes. The colored bars show the location of the genes of the CAE virus. gag encodes for the capsid protein. pol encodes a polyprotein precursor which become protease, reverse transcriptase, and integrase proteins. env encodes for both the surface and membrane components of the envelope protein. The linear order of gag, pol and env is the same in all retroviruses. vif, tat and rev code for regulatory and accessory proteins necessary for efficient viral replication including transport of transcribed proviral RNA to the cytoplasm (rev), influencing the cell cycle (tat) and protecting the viral genome from attack by cellular defenses (vif). With only a few other exceptions, these are the only genes the virus has to work with. It requires necessary proteins and enzymes found in the cell in order to replicate.
Next: the immune system and viruses